
Ретикулярная формация среднего. Вопрос47
Ретикулярной формацией называется сеть нейронов различных типов и размеров, имеющих многочисленные связи между собой, а также со всеми структурами ЦНС. Она располагается в толще серого вещества продолговатого, среднего и промежуточного мозга и регулирует уровень функциональной активности (возбудимость) всех нервных центров этих отделов ЦНС. Таким же образом она влияет на кору больших полушарий.
В ЦНС выделяют две подсистемы, выполняющие разные организующие функции: специфическую и неспецифическую . Первая обеспечивает восприятие, проведение, анализ и синтез сигналов специфической чувствительности. К ним относятся все ее виды, т.е. зрительная, слуховая, болевая и т.д.
Неспецифической подсистемой является ретикулярная формация. Она оказывает генерализованное возбуждающее или тормозящее влияние на многие структуры мозга. Следовательно, она может регулировать уровень функциональной активности моторной, сенсорной, висцеральных систем и организма в целом. Когда нервные импульсы идут по специфическим проводящим путям, по коллатералям этих путей они поступают и к нейронам ретикулярной формации. Это приводит к их диффузному возбуждению. От нейронов ретикулярной формации возбуждение передается на кору, что сопровождается возбуждением нейронов всех ее зон и слоев. Благодаря этому восходящему активирующему влиянию ретикулярной формации, повышается активность аналитико-синтетической деятельности, увеличивается скорость рефлексов, организм подготавливается к реакции на неожиданную ситуацию. Поэтому ретикулярная формация участвует в организации оборонительного, полового, пищеварительного поведения. С другой стороны, она может избирательно активировать или тормозить определенные системы мозга. В свою очередь кора больших полушарий, через нисходящие пути, может оказывать возбуждающее действие на ретикулярную формацию.
Нисходящие ретикулоспинальные пути идут от ретикулярной формации к нейронам спинного мозга. Поэтому она может оказывать нисходящие возбуждающие и тормозящие влияния на его нейроны. Например, ее гипоталамические и мезэнцефальные отделы повышают активность альфа-мотонейронов спинного мозга. В результате этого растет тонус скелетных мышц, усиливаются двигательные рефлексы. Тормозящее влияние ретикулярной формации на спинальные двигательные центры осуществляется через тормозные нейроны Реншоу. Это приводит к торможению спинальных рефлексов.
Ретикулярная формация контролирует передачу сенсорной информации через продолговатый, средний мозг, а также ядра таламуса.
Она непосредственно участвует в регуляции бодрствования и сна, за счет синхронизирующих центров сна и бодрствования, находящихся в ней.
На нейроны ретикулярной формации оказывают влияние различные фармакологические вещества: амфетамины, кофеин, LSB – 25, морфин (опыт Эдисона).
Функции мозжечка.
Мозжечок состоит их двух полушарий и червя между ними. Серое вещество образует кору и ядра. Белое образовано отростками нейронов. Мозжечок получает афферентные нервные импульсы от тактильных рецепторов, рецепторов вестибулярного аппарата, проприорецепторов мышц и сухожилий, а также двигательных зон коры. Эфферентные импульсы от мозжечка идут к красному ядру среднего мозга, ядру Дейтерса продолговатого мозга, к таламусу, а затем к мотонейроным зонам коры больших полушарий и подкорковым ядрам.
Общей функцией мозжечка является регуляция позы и движения. Эту функцию он осуществляет путем координации активности других двигательных центров: вестибулярных ядер, красного ядра, пирамидных нейронов коры. Поэтому он выполняет следующие двигательные функции:
Регуляцию мышечного тонуса позы.
Коррекцию медленных целенаправленных движений в ходе их выполнения, а также координацию этих движений с рефлексами положения тела.
Контроль за правильным выполнением быстрых движений, осуществляемых корой.
В связи с тем, что мозжечок выполняет данные функции. При его удалении у животного развивается комплекс двигательных нарушений, называемый триадой Лючиани . Он включает:
Атония и дистония – снижение и неправильное распределение тонуса скелетных мышц.
Астазия – невозможность слитного сокращения мышц, а как следствие, сохранения устойчивого положения тела при стоянии, сидении (покачивание).
Астения – быстрое утомление мышц.
Атаксия – плохая координация движений при ходьбе. Неустойчивая «пьяная» походка.
Адиадохокинез – нарушение правильной последовательности быстрых целенаправленных движений.
В клинике умеренные поражения мозжечка проявляются триадой Шарко :
Нистагм глаз в состоянии покоя.
Тремор конечностей, возникающий при их движениях.
Дизартрия – нарушение речи.
Л. А. Орбели установил, что мозжечок влияет и на различные вегетативные функции. Это влияния могут быть возбуждающими и тормозящими. Например, при раздражении мозжечка увеличивается или снижается кровяное давление, изменяется частота сердцебиений, дыхание, пищеварение. Мозжечок влияет на обмен веществ. На эти функции он воздействует через вегетативные нервные центры, координация их активности с движением. Функции внутренних органов изменяются в связи с изменением обменных процессов в них. Поэтому мозжечок оказывает на них адаптационно-трофическое влияние.
Билет 15
1. Формы (фрагменты) афферентного синтеза: Доминирующая мотивация; Обстановочная афферентация; Пусковая афферентация. Роль ретикулярной формации.
2. Быстрые и медленные мышечные волокна.
Вопрос 1
АФФЕРЕНТНЫЙ СИНТЕЗ - (соединение, составление) - процесс сопоставления, отбора и синтеза многочисленных и разных по функциональному значению афферентаций, вызванных различными воздействиями на организм, происходящий в ц. н. с., на основе к-рого формируется цель действия.
А. с. согласно теории функциональной системы Анохина - первая, универсальная, стадия любого целенаправленного поведенческого акта (см. Функциональные системы).
А. с. включает обработку 4 основных видов афферентных возбуждений.
1. Мотивационное возбуждение отражает доминирующую потребность организма, к-рая возникает под влиянием метаболических, гормональных, а у человека - и социальных факторов Мотивация играет решающую роль в формировании цели действия. Специфически повышая реактивность корковых нейронов с помощью ориентировочно-исследовательской реакции , мотивационное возбуждение способствует обработке и активному отбору сенсорной информации, необходимой для построения целенаправленного поведения.
2. Обстановочная афферентация представляет собой воздействие на организм всей совокупности внешних факторов, составляющих конкретную обстановку, на фоне к-рой развертывается приспособительная деятельность. Обстановочная афферентация формируется не только постоянными компонентами обстановки, но и рядом последовательных афферентных воздействий на организм. Характерная особенность обстановочной афферентации состоит в том, что она придает специфику будущей поведенческой реакции, обеспечивая ее приспособительное значение только в данной обстановке.
Наиболее отчетливо роль обстановочной афферентации проявляется в опытах с условными рефлексами. В этих случаях на один и тот же условный раздражитель животное отвечает условной оборонительной реакцией в одной экспериментальной камере и условной пищевой - в другой (или в одной и той же экспериментальной камере утром животное отвечает пищевой реакцией, а вечером - оборонительной).
На стадии афферентного синтеза решаются вопросы “что делать?”, “как делать?”, “когда делать?”.
Пусковая афферентация
Представляет собой специальный раздражитель, собственно запускающий поведенческую реакцию. Значение пускового раздражителя состоит в том, что он призван обозначить момент начала поведенческой реакции.
Целенаправленное поведение может начинаться и без явного пускового стимула. Примерами таких реакций являются регулярно совершающиеся физиологические отправления (еда, сон, дефекация, мочеиспускание и др.), приуроченные к определенным периодам суток.
Афферентный синтез осуществляется на основе следующих нейрофизиологических механизмов:
1) механизмы восходящих активирующих влияний подкорковых образований на кору головного мозга. Это прежде всего, активирующие влияния гипоталамуса к лобным отделам коры, через передние ядра таламуса, что отражает мотивационные возбуждения. Аналогичным образом воздействуют другие лимбические системы. Вторыми по активирующему значению являются ретикулярные структуры среднего мозга и моста, которые обеспечивают соответствующий уровень бодрствования.
2) механизмы конвергенции возбуждений различного качества на нейронах коры и подкорковых структурах мозга. В частности, мультисенсорная конвергенция от поверхностей (зрительная, тактильная, слуховая, температурная и др.); мультибиологическая конвергенция, связанная с определенными состояниями (голод, боль и т.п.) и др.;
3) интеграцию мотивационных, обстановочных и пусковых афферентаций на нейронах коры мозга;
4) механизмы формирования доминанты, за счет которых подавляется текущая деятельность и удерживается вновь сформированная поведенческая реакция.
Роль ретикулярной формации
Ретикулярная формация характеризуется относительно малой возбудимостью. Эффекты ее раздражения появляются через большой латентный период, она медленно реагирует и остается активной в течение продолжительного времени после прекращения раздражения (длительное последействие). Ретикулярная формация облегчает или подавляет фазные движения и напряжение скелетных мышц, вызываемые мотонейронами спинного мозга, а также движения, вызванные с коры больших полушарий. Ретикулярная формация среднего и промежуточного мозга облегчает рефлекторные движения животных, раздражение промежуточного мозга тормозит двигательные рефлексы спинного мозга.
Боковые отделы ретикулярной формации варолиева моста и среднего мозга облегчают, а средние ее отделы в продолговатом мозге тормозят двигательные рефлексы. Облегчение и торможение зависят также от интенсивности и продолжительности раздражения ретикулярной формации. По гамма-нейронам она регулирует функции мышечных веретен, следовательно, обратную информацию из скелетных мышц. Она изменяет также возбудимость восходящих афферентных путей спинного мозга, что может снизить или прекратить постсинаптическое торможение. Тонические влияния ретикулярной формации вызывают ВПСП или ТПСП в мотонейронах спинного мозга. Она изменяет также передачу импульсов в мозговом стволе и одновременно с влиянием на скелетную мускулатуру вызывает сосудодвигательные, дыхательные, зрачковые и другие реакции.
Ретикулярная формация оказывает адаптационно-трофическое влияние на кору больших полушарий, подкорковые образования промежуточного мозга, мозжечок и спинной мозг. Существуют взаимные влияния этих отделов нервной системы, как возбуждающие, так и тормозящие. Она участвует в физиологических процессах сна и пробуждения, а также в эмоциях, в реакции напряжения («стресс») и др. Раздражение ретикулярной формации вызывает пробуждение спящих животных, а ее разрушение и выключение - глубокий сон у бодрствующих животных. Изучены взаимные влияния ретикулярной формации и коры больших полушарий.Установлено участие ретикулярной формации в образовании и протекании условных рефлексов
По симпатическим волокнам ретикулярная формация регулирует возбудимость и работоспособность скелетной мускулатуры, функциональное состояние нервной системы и органов чувств, оказывая на них адаптационно-трофическое влияние. Регуляция рефлексов позы и двигательных рефлексов, перемещающих тело, осуществляется по эфферентным гамма-волокнам иннервирующим проприоцепторы.
Ретикулярная формация регулирует вегетативные функции, деятельность внутренних органов. Она влияет на образование гормонов в гипофизе и других железах внутренней секреции и в ней концентрируются гормоны и медиаторы.
Афферентные волокна поступают в нее по симпатическим и блуждающим нервам. Частьклеток ретикулярной формации среднего мозга и варолнева моста возбуждается адреналином и норадреналином (адренореактивные системы) а другая часть, расположенная в промежуточном мозге, несколько выше среднего мозга, возбуждается ацетилхолином и его производными (холинореактивные системы). Адренореактивные системы среднего мозга и варолиева моста облегчают наступление двигательных рефлексов, а адренореактивные системы продолговатого мозга тормозят спинномозговые рефлексы. Адреналин возбуждает и холинореактивные системы. Предполагается, что действие ацетилхолина и его производных менее ограничено, чем действие адреналина, и охватывает многие области головного мозга. Действие ацетилхолина на ретикулярную формацию противоположно его периферическому влиянию на внутренние органы. Ретикулярную формацию среднего и продолговатого мозга возбуждает углекислота.
Гормоны и медиаторы действуют на функцию больших полушарий как непосредственно, так и посредством ретикулярной формации. Таким образом, ретикулярная формация мозгового ствола - подкорковый центр вегетативной нервной системы.
Вопрос 2 .
– ряд ядерных структур, занимающих центральное место в стволовой части мозга. Морфологическое строение ретикулярной формации очень напоминает сеточку, и именно на этот признак в первую очередь обратил внимание немецкий анатом Отто Дейтерс. Именно по этому признаку он и назвал эту структуру (лат. reticulum - сеточка, formatio - образование). Согласен! «Сетчатое образование» — звучит не так круто 🙂
Так как ретикулярная формации проходит через весь ствол головного мозга, то анатомически её можно было бы разделить на отделы продолговатого мозга, варолиева моста и среднего мозга, но т. к. отдельные части этой структуры заняты общим для них делом, то ее рассматривают, как единую структуру.
Для понимания приведу сравнение… Если вы когда-нибудь бывали на концертах или хотя бы смотрели их по телевизору, то наверняка замечали огромные панели с кучей кнопочек, тумблеров, переключателей и т. д. С помощью этих панелей оператор настраивает качество звука, приглушая одни частоты и повышая другие, а так же цветность, яркость, контраст и т. д. Так вот ретикулярная формация занимается именно этим. Т. е. она получает сигналы абсолютно от всех нисходящих и восходящих путей, перерабатывает их, вырабатывает новые координирующие сигналы и выдает их по назначению, обеспечивая нам нормальное восприятие.
Некоторые нейроны Р. ф. проявляют фоновую активность, разряжаясь 5-10 раз в секунду. Эти нервные центры оказывают влияния на кору головного мозга, постоянно поддерживая в нас сознание. При разрушении этих центров у животных наступает .
Для сравнения поясню. Ретикулярная формация поддерживает в нас сознание так же, как огонь в костре поддерживает кипение воды в котле. Стоит потушить огонь — и кипящая вода, так же как и сознание, перестает проявлять активность. Следовательно, в Р. ф. находятся одни из центров сна и бодрствования.
С корой головного мозга вообще связи особенные. Понятно, что Р.Ф. отвечает за и возбуждения в коре мозга. Кора мозга, в свою очередь, также оказывает тормозные и возбуждающие влияния на ретикулярную формацию. Образуя замкнутые нейронные связи, эти две системы взаимно регулируют друг друга и уравновешивают свои влияния.
Выводы
Функции ретикулярной формации изучены не достаточно ввиду высокой степени сложности организации данной структуры, но имеющихся данных достаточно для следующих выводов:
Оказывает влияние на уровень сознания путем взаимодействия в кору мозга. Участвует в цикле в придание эмоциональной окраски сенсорным сигналам в т. ч. болевым, путем проведения афферентной информации к лимбической системе. Путем взаимного координирования афферентных и эфферентных систем участвует в формировании жизненно важных рефлексов. Также принимает участие в вегетативных функциях организма и как важная составляющая двигательных центров ствола мозга.
Сложное строение человеческого мозга открывает тайны нашего поведения, объясняет законы мыслительной деятельности, протекание эмоций и чувств. Каждое полушарие головного мозга отвечает за свои специфические функции и задачи (например, известно, что правое отвечает за логику, а левое - за воображение и фантазию), но имеются также структуры, которые обеспечивают единую и согласованную работу всей центральной нервной системы. Одной из таких структур является ретикулярная формация.
Общие сведения
Ретикулярная формация - это участок стволового отдела мозга, представленный разветвленной сетью нервных клеток и ядер, соединяющих между собой различные отделы головного мозга. В отличие от других структур, - например, таламуса, гипоталамуса, мозжечка, - которые имеют некую цельную форму (ядра, железы), ретикулярная формация не представлена единым морфологическим образованием, а является «сетью» (от латинского reticulum — сеть) дендритов и аксонов, которые с разной степенью плотности проникают между отделами и структурами головного мозга, объединяя их между собой и обеспечивая их совместную деятельность.
Метафорически выражаясь: если наш мозг представить в виде некоего изделия, скажем, рубашки, то ретикулярная формация - это нити, которыми рубашка сшита. Ретикулярная формация пронизывает структуры продолговатого, среднего мозга и моста, имеет непосредственные связи с мозжечком, спинным мозгом, таламусом и опосредованные - с вышележащими отделами: гипоталамусом, зрительными ядрами и корой.
Как устроена
В состав ретикулярной формации входит огромное количество нейронов с разветвленными дендритами и длинными аксонами, за счет чего становится возможным передавать нервные импульсы в различные отделы головного и спинного мозга. При этом можно выделить две наиболее крупные группы нейронных скоплений:
- Ретикулотегментальное ядро, нейроны которого получают сигналы от вышележащих отделов ГМ (четверохолмие, таламус) и передают их далее в структуры мозжечка, регулируя тем самым некоторые жизненно важные двигательные функции: координация взгляда, движения глаз.
- Латеральное ядро, нейроны которого восходят от структур спинного мозга и вестибулярных ядер и обеспечивают информирование коры ГМ о положении тела в пространстве, участвуют в регуляции дыхания и сосудистой иннервации.
- Кроме того, в состав ретикулярной формации входят нейроны, которые принимают важное участие в работе центров терморегуляции, насыщения и голода.

Основные функции
Основное предназначение ретикулярной формации состоит в сенсорном анализе многочисленных сигналов, поступающих от различных отделов ГМ.
За счет тесных связей со спинным мозгом она также принимает самое активное участие в моторной регуляции, начиная от глотательного рефлекса и заканчивая сложными двигательными операциями. Кроме того, ретикулярная формация осуществляет активирующее воздействие на весь ГМ в целом, участвуя в регуляции циклов сна и бодрствования.
В общем виде функции ретикулярной формации сводятся к следующим:
- Регуляция скелетной мускулатуры (участвует в управлении движениями тела) и вегетативных функций (дыхание, чихание, кровообращение и т.п.).
- Контроль процессов сна и бодрствования (за счет оказания активирующего и тормозящего воздействия на кору больших полушарий).
- Активизирующая функция (проявляется в том, что ретикулярная формация обеспечивает постоянное тоническое возбуждение коры ГМ, за счет чего становится возможным поддержание внимания, сознания и протекание мыслительных процессов.)
- Обработка сигналов, поступающих из внешней и внутренней среды.
Отличительные особенности работы ретикулярной формации связаны, прежде всего, с определенными свойствами ее :
Нервные клетки, входящие в состав ретикулярной формации, обладают повышенной способностью к тоническому возбуждению. Это означает, что бо́льшая часть нейронов находятся в постоянном возбуждении и генерирует нервные импульсы, которые передаются в вышележащие отделы ГМ. Такая тоническая активность обусловлена рядом факторов:
- Проходимость большого количества сигналов через структуры ретикулярной формации. Приведем простую аналогию: представьте себе рояль или какой-либо другой струнный инструмент. Понятно, что когда мы напрямую касаемся струн, они начинают колебаться и издавать звук. То же самое происходит и с нервными клетками, когда к ним приходят сигналы от других нейронов.Однако представим далее, что мы не касаемся непосредственно струн инструмента, но, скажем, скачем рядом с ним, сильно стуча ногами о пол. Звука инструмента мы, может быть, и не услышим, но едва заметное колебание струн все же будет иметь место. То же самое происходит и с нейронами ретикулярной формации. Поскольку через нее постоянно проходят какие-то сигналы (как афферентные, так и эфферентные) от различных структур ЦНС, то это и создает постоянное тоническое возбуждение нейронов ретикулярной формации, за счет того, что она находится в эпицентре постоянного нервно-импульсного обмена.
- Повышенная чувствительность нейронов к химическим веществам (гормоны, медикаменты, психотропные вещества). Чашка кофе, выпитая утром, «включает» именно структуры ретикулярной формации и за счет длительного сохранения возбуждения в ее нейронах поддерживает нас в активном состоянии.
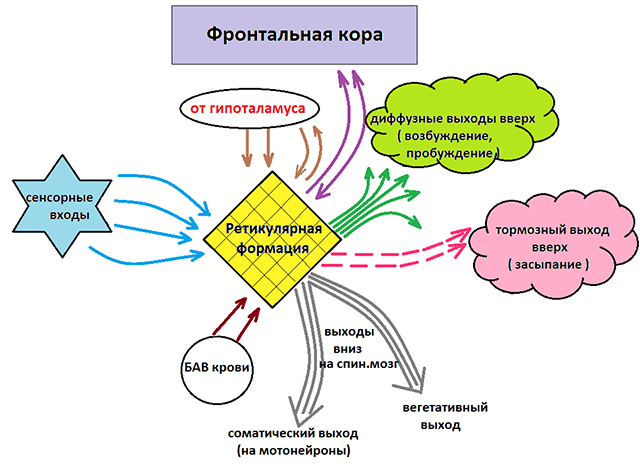
Нисходящее и восходящее влияние РФ
Как уже отмечалось, ретикулярная формация оказывает возбуждающее и тормозящее действие на различные отделы ГМ. При этом можно выделить два отдела, которые специализируются на передаче возбуждения в те или иные структуры мозга.
Нисходящий отдел: представлен вегетативными и двигательными центрами и оказывает нисходящее влияние на отделы спинного мозга. Соответствующие нейронные скопления регулируют деятельность дыхательного, сосудодвигательного, слюноотделительного центров, а также центров, отвечающих за построение простых и сложных двигательных реакций. Это указывает на решающую роль именно центральной нервной системы в регуляции даже элементарных безусловных рефлексов. Стимуляция нисходящего отдела приводит к затормаживанию спинальных центров и вызывает в естественной среде состояние глубоко сна (спать «без задних ног»). Тот же эффект можно вызвать и искусственным путем, например, вводя человека в состояние транса или наркоза.
Восходящий отдел: представлен нервными волокнами, соединяющими структуры ретикулярной формации с вышележащими отделами: таламусом, гипоталамусом, мозжечком и корой. Восходящее влияние оказывает стимулирующее действие на кортикальные структуры и обеспечивает активное состояние сознания. Восходящее влияние не прекращается, даже когда мы спим. Если бы наш мозг мог полностью «отключаться», то каждое пробуждение было бы сродни рождению: кто я? Где я? Как я сюда попал? Однако за счет работы ретикулярных структур у нас сохраняется возможность всегда возвращаться в то исходное состояние сознания, в котором мы находились до момента сна. Кроме того, и во время ночного отдыха у нас остается способность реагировать на некоторые жизненно важные раздражители, т.е. мы, как правило, не спим «мертвым сном» и можем проснуться, если рядом зашевелился и заплакал ребенок, что-то громко упало и т.п.
Проявление повреждения структур
Ретикулярная формация играет значимую роль в интегративной деятельности всего головного мозга. За счет выполнения функции ведущего проводника всех видов нервных импульсов во все отделы ЦНС, ретикулярная формация находится в постоянной работе. Чрезмерные умственные и эмоциональные перегрузки вредны для мозга в целом и для ретикулярной формации, в частности. К счастью, своевременный прием седативных препаратов может (за счет повышенной восприимчивости нейронов к химическому воздействию) быстро исправить ситуацию и нормализовать состояние.
Однако возможны и менее благоприятные исходы. Повреждение возможно в результате черепно-мозговых травм, онкологических заболеваний головного мозга, инфекционных поражений.
Основным проявлением неблагополучия является потеря сознания.
Нарушение восходящих связей обнаруживает себя в состоянии апатии, слабости, повышенной сонливости, двигательной растороможенности, нарушениях ночного сна. Нередки сопутствующие вегетативные расстройства.
Ретикулярная формация (лат. rete - сеть) представляет собой совокупность клеток, клеточных скоплений и нервных волокон, расположенных на всем протяжении ствола мозга (продолговатый мозг, мост, средний и промежуточный мозг) и в центральных отделах спинного мозга. Ретикулярная формация получает информацию от всех органов чувств, внутренних и других органов, оценивает ее, фильтрует и передает в лимбическую систему и кору большого мозга. Она регулирует уровень возбудимости и тонуса различных отделов центральной нервной системы, включая кору большого мозга, играет важную роль в сознании, мышлении, памяти, восприятии, эмоциях, сне, бодрствовании, вегетативных функциях, целенаправленных движениях, а также в механизмах формирования целостных реакций организма. Ретикулярная формация прежде всего выполняет функцию фильтра, который позволяет важным для организма сенсорным сигналам активировать кору мозга, но не пропускает привычные для него или повторяющиеся сигналы.
Ретикулярная формация представляет собой важный пункт на пути восходящей неспецифической соматосенсорной системы. Соматовисцеральные афференты идут в составе спиноретикулярного тракта (переднебоковой канатик), а также, возможно, в составе проприоспинальных (полисинаптических) путей и соответствующих путей от ядра спинального тройничного тракта. К ретикулярной формации приходят также пути от всех других афферентных черепномозговых нервов, т.е. практически от всех органов чувств. Дополнительная афферентация поступает от многих других отделов головного мозга - от моторных областей коры и сенсорных областей коры, от таламуса и гипоталамуса. Имеется также множество эфферентных связей - нисходящие к спинному мозгу и восходящие через неспецифические таламические ядра к коре головного мозга, гипоталамусу и лимбической системе. Большинство нейронов образует синапсы с двумя - тремя афферентами разного происхождения, такая полисенсорная конвергенция характерна для нейронов ретикулярной формации. Другими их свойствами являются большие рецептивные поля поверхности тела, часто билатеральные, длительный латентный период ответа на периферическую стимуляцию (вследствие мультисинаптического проведения), слабая воспроизводимость реакции (стохастические колебания числа потенциалов действия при повторной стимуляции). Все эти свойства противоположны свойствам лемнисковых нейронов в специфических ядрах соматосенсорной системы (рис.9-7 и рис. 5-13).
Функции ретикулярной формации изучены не полностью. Считается, что она участвует в следующих процессах:
1. в регуляции уровня сознания путем воздействия на активность корковых нейронов, например, участие в цикле сон / бодрствование,
2. в придании аффективно-эмоциональной окраски сенсорным стимулам, в том числе болевым сигналам, идущим по переднебоковому канатику, путем проведения афферентной информации к лимбической системе,
3. в вегетативных регулирующих функциях, в том числе во многих жизненно важных рефлексах (циркуляторных рефлексах и дыхательных рефлексах, рефлекторных актах глотания, кашля, чихания), при которых должны взаимно координироваться разные афферентные и эфферентные системы,
4. в целенаправленных движениях в качестве важного компонента двигательных центров ствола мозга.

Вопрс48. Сравнительная характеристика кабельного и сальтаторного видов проведения возбуждения
Нервная ткань обладает таким физиологическим свойством как проводимость, т. е. способностью проводить возбуждение по ходу нервного волокна в виде потенциала действия. Выделяют два вида проведения возбуждения в зависимости от строения нервного волокна. Различают два вида нервных волокон: мякотные (миелиновые) и безмякотные (немиелиновые). В безмякотных нервных волокнах наблюдается непрерывное распространение возбуждения, в основе которого лежат локальные или круговые токи. Как было сказано раньше, возбужденный электроотрицательный участок нервного волокна становится раздражителем для ближайшего невозбужденного электроположительного участка, который возбуждается (возбужденный участок как бы разряжается в сторону невозбужденного, следствием чего и являются появление локальных или круговых токов).
Миелин, прокрывающий нервное волокно, располагается сегментами, т. е. прерывисто. Миелин - хороший изолятор и, если бы он сплошным слоем покрывал нервное волокно, то возбуждение не распространялось бы. Миелиновая оболочка образуется клетками неврилеммы или шванновскими клетками. Плазматическая мембрана одной шванновской клетки обвертывает спирально в несколько слоев участок аксона, длиной в несколько сотых микрон. Между участками аксона, покрытого миелиновой оболочкой, остаются немиеленизированные зоны. Эти зоны называются перехватами Ранвье.
В волокнах, покрытых миелиновой оболочкой (мякотные волокна) возбуждение распространяется скачкообразно (сальтаторно), т. е. по перехватам Ранвье. Как было показано японским физиологом Тасаки, это создает своеобразную систему надежности для распространения возбуждения (разности потенциалов между возбужденным и невозбужденным участками волокна хватает на 5-6 перехватов Ранвье). В случае, если появится повреждение на небольшом участке волокна вследствие большого электрического поля распространение потенциала действия не нарушается. Как известно, начальная часть аксона в нервной клетке не покрыта миелиновой оболочкой. Именно в этом участке волокна и формируется потенциал действия. Возникает разность потенциалов между возбужденным и невозбужденным участком первого перехвата Ранвье, который под влиянием этого электрического поля возбуждается. Затем разность потенциалов формируется между возбужденным первым перехватом Ранвье и следующим, который перезаряжается и возбуждение приобретает распространяющий характер. Таким образом, в основе распространения возбуждения по мякотному волокну, как и безмякотному, лежат также местные (круговые, вихревые) токи. В перехватах Ранвье, находящихся друг от друга на расстоянии 2 мм, обнаружена большая плотность натриевых каналов - до 1200 на 1 мкм2, что значительно облегчает проведение возбуждения по нервному волокну. Прерывистое распространение возбуждения имеет некоторые преимущества по сравнению с непрерывным. Во-первых, скорость распространения возбуждения в волокнах, покрытых миелином, в 8-10 раз быстрее, чем в безмякотных. Во-вторых, на распространение возбуждения прерывистого типа затрачивается меньше энергии, оно более экономично, что, по всей вероятности, связано с большой плотностью натриевых каналов в перехватах Ранвье.
При распространении возбуждения по нервному волокну следует учитывать чисто физические или кабельные свойства проводника (нерв можно представить как кабель, помещенный в морскую воду). К кабельным свойствам относится, в частности, диаметр (поперечное сечение) проводника - чем толще нервное волокно (или больше поперечное сечение), тем меньше сопротивление. Следовательно, тем быстрее будет распространение возбуждения в виде импульса. Большое значение при возбуждении имеет также емкость и сопротивление мембраны. Так, если входное сопротивление мембраны больше, то и возбудимость в этом месте уменьшается. К кабельным свойствам относится также и электротон, оказывающий большое влияние на проводимость: чем выраженнее катэлектротон, тем быстрее проводится потенциал действия. Анэлектротонические изменения, напротив, ухудшают проведение возбуждения по нервной ткани.
В зависимости от скорости проведения возбуждения все нервные волокна делятся на три группы: А, В и С. Нервные волокна группы А - это высокоскоростные волокна, исключительно мякотного типа. В зависти от сечения нервного волокна скорость проведения возбуждения их колеблется в пределах 20-120 м/с. Различают А- волокна - самые скоростные - 70-120 м/с (диаметр волокна 12-20 мкм - a-волокна, их средняя скорость проведения возбуждения составляет 70-120 м/с; диаметр 8-12 мкм - b-волокна, проводящие возбуждение со скоростью 40-70 м/с; диаметр волокна 4-8 мкм - g-волокна, проводящие возбуждение со скоростью 20-40 м/с). Таким образом, чем толще проводник, тем больше скорость проведения возбуждения. Нервные волокна группы В представляют собой в основном безмякотные волокна, скорость распространения возбуждения которых составляет 6-20 м/с. Нервные волокна группы С представлены исключительно безмякотными волокнами вегетативной природы, скорость проведения возбуждения их составляет 0,5-6 м/с.
В физиологии имеется три закона распространения возбуждения.
Закон целостности нерва (закон непрерывности). Нерв проводит возбуждение только в том случае, если он сохраняет свою гистологическую и функциональную целостность. Любые отклонения этих показателей приводят к нарушению его проводимости. Действие местных анестетиков (новокаин) основано на том, что молекулы новокаина блокируют натриевые каналы, в результате чего прекращается натриевый ток и ткань теряет способность возбуждаться. Возбуждение при раздражении болевых рецепторов доходит до места, где действует новокаин и блокируется, вследствие чего болевые импульсы не достигают болевого центра.
Закон двустороннего проведения возбуждения. Нервное волокно способно проводить возбуждение от рецепторов к центрам и наоборот, от центров к периферическим образованиям. Такая закономерность была показана в классических исследования Кюне и Бабухина. Так, опыт Кюне заключался в следующем: если нарушить целостность мышцы между двумя ее участками, которые иннервируются двумя разветвлениями одного аксона, то электрическое раздражение любого из ответвлений аксона приводит к сокращению обеих частей мышцы.
Закон изолированного распространения возбуждения. Известно, что потенциал действия в волокнах, покрытых миелином, не перебрасывается с одного нервного волокна на другое благодаря хорошим изоляционным свойствам миелина. Такое изолированное проведение возбуждения обеспечивает мелкие и точные профессиональные сокращения мышц (игра на пианино, работа часового мастера и др.). Сразу после рождения достаточная миелинизация нервных волокон отсутствует и на любое раздражение новорожденные в большинстве случаев отвечают не локальными, а диффузными сокращениями большой группы мышц. Подобная же ответная реакция наблюдается во всех гладких мышцах, которые иннервируются безмякотными нервными волокнами, не обладающими изоляционнымитсвойствами.