
Retikulární formace středu. Otázka 47
Retikulární formace Nazývá se síť neuronů různých typů a velikostí, které mají četná spojení mezi sebou, stejně jako se všemi strukturami centrálního nervového systému. Nachází se v tloušťce šedé hmoty prodloužené míchy, středního mozku a diencefala a reguluje úroveň funkční aktivity (vzrušivosti) všech nervových center těchto částí centrálního nervového systému. Stejně tak působí na mozkovou kůru.
V CNS se rozlišují dva subsystémy, které plní různé organizační funkce: charakteristický a nespecifické. První poskytuje vnímání, vedení, analýzu a syntézu signálů specifické citlivosti. Patří mezi ně všechny její druhy, tzn. zrakový, sluchový, bolestivý atd.
nespecifické subsystémem je retikulární formace. Má generalizovaný excitační nebo inhibiční účinek na mnoho mozkových struktur. V důsledku toho může regulovat úroveň funkční aktivity motorického, senzorického, viscerálního systému a těla jako celku. Když nervové impulsy putují po specifických drahách, dostávají se také do neuronů retikulární formace podél kolaterál těchto drah. To vede k jejich difúznímu buzení. Z neuronů retikulární formace se přenáší vzruch do kortexu, který je doprovázen buzením neuronů ve všech jeho zónách a vrstvách. Díky tomuto vzestupně aktivačnímu vlivu retikulární formace se zvyšuje aktivita analytické a syntetické aktivity, zvyšuje se rychlost reflexů, tělo se připravuje na reakci na neočekávanou situaci. Proto se retikulární formace podílí na organizaci obranného, sexuálního a trávicího chování. Na druhou stranu může selektivně aktivovat nebo inhibovat určité mozkové systémy. Na druhé straně kůra hemisféry, prostřednictvím sestupných drah, může mít stimulační účinek na retikulární formaci.
Sestupné retikulospinální dráhy jdou od retikulární formace k neuronům míchy. Proto může uplatňovat sestupné excitační a inhibiční vlivy na své neurony. Jeho hypotalamický a mezencefalický úsek například zvyšují aktivitu míšních alfa motorických neuronů. V důsledku toho se zvyšuje tonus kosterních svalů, zvyšují se motorické reflexy. Inhibiční účinek retikulární formace na spinální motorická centra se provádí prostřednictvím Renshawových inhibičních neuronů. To vede k inhibici míšních reflexů.
Retikulární formace řídí přenos smyslových informací přes prodlouženou míchu, střední mozek a jádra thalamu.
Přímo se podílí na regulaci bdění a spánku, a to díky synchronizačním centrům spánku a bdění v něm umístěných.
Na neurony retikulární formace působí různé farmakologické látky: amfetaminy, kofein, LSB - 25, morfin (Edisonův experiment).
Funkce mozečku.
Mozeček se skládá ze dvou hemisfér a vermis mezi nimi. Šedá hmota tvoří kůru a jádra. Bílá je tvořena procesy neuronů. Mozeček přijímá aferentní nervové impulsy z hmatových receptorů, receptorů vestibulárního aparátu, proprioceptorů svalů a šlach a také motorických oblastí kůry. Eferentní impulsy z mozečku jdou do červeného jádra středního mozku, Deitersova jádra prodloužené míchy, do thalamu a poté do motoneuronových zón mozkové kůry a subkortikálních jader.
Obecnou funkcí mozečku je regulace držení těla a pohybu. Tuto funkci plní koordinací činnosti dalších motorických center: vestibulárních jader, červeného jádra, pyramidálních neuronů kůry. Proto provádí následující motorické funkce:
nařízení svalový tonus představuje.
Korekce pomalých účelových pohybů při jejich provádění a také koordinace těchto pohybů s reflexy polohy těla.
Kontrola nad správným prováděním rychlých pohybů prováděných kůrou.
Vzhledem k tomu, že tyto funkce plní mozeček. Při jeho odstranění se u zvířete vyvine komplex motorických poruch tzv Lucianiho triáda . To zahrnuje:
Atonie a dystonie- Snížené a nesprávné rozložení tonusu kosterního svalstva.
astasie- nemožnost kontinuální svalové kontrakce a v důsledku toho udržení stabilní polohy těla při stání, sezení (houpání).
astenie- rychlá svalová únava.
Ataxie- špatná koordinace pohybů při chůzi. Nejistá "opilá" chůze.
Adiadochokineze- porušení správného sledu rychlých účelných pohybů.
Na klinice se projevují středně těžké léze mozečku Charcotova triáda:
Oční nystagmus v klidu.
Třes končetin, ke kterému dochází při jejich pohybech.
Dysartrie je porucha řeči.
L. A. Orbeli zjistil, že mozeček ovlivňuje i různé autonomní funkce. Tyto vlivy mohou být excitační nebo inhibiční. Když je například stimulován mozeček, krevní tlak se zvyšuje nebo snižuje, srdeční frekvence, dýchání a trávení se mění. Mozeček ovlivňuje metabolismus. Ovlivňuje tyto funkce prostřednictvím autonomních nervových center, koordinuje jejich činnost s pohybem. Funkce vnitřních orgánů se mění v důsledku změn metabolických procesů v nich. Proto na ně mozeček působí adaptačně-troficky.
Vstupenka 15
1. Formy (fragmenty) aferentní syntézy: Dominantní motivace; situační aferentace; Spuštění aferentace. Role retikulární formace.
2. Rychlá a pomalá svalová vlákna.
Otázka 1
AFERENTNÍ SYNTÉZA- (spojení, kompilace) - proces porovnávání, výběru a syntézy četných a funkčních významností různých aferentací způsobených různými účinky na organismus, vyskytující se v c. n. s., na jejímž základě se tvoří účel žaloby.
Tak jako. podle Anokhinovy teorie funkčního systému je to první, univerzální, stádium jakéhokoli účelového behaviorálního aktu (viz Funkční systémy).
Tak jako. zahrnuje zpracování 4 hlavních typů aferentních vzruchů.
1. Motivační excitace odráží dominantní potřebu těla, která vzniká vlivem metabolických, hormonálních a u člověka - sociálních faktorů Motivace hraje rozhodující roli při utváření cíle jednání. Konkrétně zvýšení reaktivity kortikálních neuronů s pomocí průzkumná reakce, motivační vzrušení přispívá ke zpracování a aktivnímu výběru smyslových informací nezbytných pro konstrukci cíleného chování.
2. Situační aferentace je působení na tělo celého agregátu vnější faktory, které tvoří konkrétní situaci, na jejímž pozadí se odvíjí adaptivní činnost. Situační aferentaci tvoří nejen konstantní složky situace, ale i řada po sobě jdoucích aferentních vlivů na organismus. Vlastnosti situační aferentace spočívá v tom, že dává specifičnost budoucí behaviorální reakci a poskytuje její adaptivní hodnotu pouze v dané situaci.
Role situační aferentace se nejzřetelněji projevuje v experimentech s podmíněnými reflexy. V těchto případech zvíře reaguje na stejný podmíněný podnět podmíněnou obrannou reakcí v jedné pokusné komoře a podmíněnou potravní reakcí ve druhé (nebo ve stejné pokusné komoře ráno zvíře reaguje potravní reakcí a v večer s obrannou).
Ve fázi aferentní syntézy se řeší otázky „co dělat?“, „jak dělat?“, „kdy dělat?“.
Spuštění aferentace
Je to zvláštní podnět, který ve skutečnosti spouští behaviorální reakci. Význam startovacího podnětu spočívá v tom, že má indikovat okamžik nástupu behaviorální reakce.
Chování zaměřené na cíl může začít bez explicitního spouštěcího podnětu. Příklady takových reakcí jsou pravidelné fyziologické funkce (jídlo, spánek, defekace, močení atd.) určitá období dní.
Aferentní syntéza se provádí na základě následujících neurofyziologických mechanismů:
1) mechanismy vzestupně aktivačních vlivů podkorových útvarů na mozkovou kůru. Především je to aktivační působení hypotalamu na frontální kortex, prostřednictvím předních jader thalamu, které odráží motivační vzruchy. Ostatní limbické systémy působí podobně. Druhou aktivační hodnotou jsou retikulární struktury středního mozku a mostu, které poskytují odpovídající úroveň bdělosti.
2) mechanismy konvergence vzruchů různé kvality na neuronech kůry a subkortikálních strukturách mozku. Zejména multisenzorická konvergence z povrchů (zrakových, hmatových, sluchových, teplotních atd.); multibiologická konvergence spojená s určitými stavy (hlad, bolest atd.) atd.;
3) integrace motivačních, situačních a spouštěcích aferentací na neuronech mozkové kůry;
4) mechanismy utváření dominanty, díky nimž je současná aktivita potlačena a nově vytvořená behaviorální reakce je zachována.
Role retikulární formace
Retikulární formace vyznačující se relativně nízkou excitabilitou. Účinky jejího podráždění se objevují po dlouhém latentním období, reaguje pomalu a zůstává aktivní ještě dlouho po odeznění podráždění (long aftereffect). Retikulární formace usnadňuje nebo potlačuje fázické pohyby a napětí kosterního svalstva způsobené motorickými neurony míchy a také pohyby způsobené z mozkové kůry. Retikulární formace středního mozku a diencefala usnadňuje reflexní pohyby zvířat, dráždění diencephala inhibuje motorické reflexy míchy.
Boční úseky retikulární formace mostu a středního mozku usnadňují a jeho střední úseky v prodloužené míše inhibují motorické reflexy. Úleva a inhibice závisí také na intenzitě a trvání stimulace retikulární formace. Prostřednictvím gama neuronů reguluje funkce svalových vřetének, a tedy zpětnou vazbu od kosterních svalů. Mění také excitabilitu vzestupných aferentních drah míchy, což může snížit nebo zastavit postsynaptickou inhibici. Tonické vlivy retikulární formace způsobují EPSP nebo IPSP v motorických neuronech míchy. Mění také přenos vzruchů v mozkovém kmeni a současně s působením na kosterní svaly vyvolává vazomotorické, respirační, zornicové a další reakce.
Retikulární formace působí adaptačně-troficky na mozkovou kůru, subkortikální formace diencephalon, mozeček a míchu. Dochází k vzájemnému ovlivňování těchto útvarů nervový systém jak excitační, tak inhibiční. Podílí se na fyziologických procesech spánku a probouzení, dále na emocích, na stresové reakci („stres“) atd. Podráždění retikulární formace způsobuje probuzení spících zvířat a její zničení a vypnutí způsobuje hluboký spánek u bdělých zvířat. Byly studovány vzájemné vlivy retikulární formace a mozkové kůry a byla stanovena účast retikulární formace na vzniku a průběhu podmíněných reflexů.
Prostřednictvím sympatických vláken reguluje retikulární formace dráždivost a výkonnost kosterních svalů, funkční stav nervového systému a smyslových orgánů a působí na ně adaptačně-troficky. Regulace reflexů držení těla a motorických reflexů, které pohybují tělem, se provádí podél eferentních gama vláken inervujících proprioreceptory.
Retikulární formace reguluje autonomní funkce, činnost vnitřních orgánů. Ovlivňuje tvorbu hormonů v hypofýze a dalších žlázách s vnitřní sekrecí a koncentrují se v ní hormony a mediátory.
Aferentní vlákna do něj vstupují prostřednictvím sympatického a vagusového nervu. Část buněk retikulární formace středního mozku a varolneválního můstku je excitována adrenalinem a norepinefrinem (adrenergní systémy) a druhá část, umístěná v diencefalu, mírně nad středním mozkem, je excitována acetylcholinem a jeho deriváty (cholinoreaktivní systémy ). Adrenoreaktivní systémy středního mozku a mostu usnadňují nástup motorických reflexů a adrenoreaktivní systémy prodloužené míchy inhibují míšní reflexy. Adrenalin také stimuluje cholinergní systémy. Předpokládá se, že působení acetylcholinu a jeho derivátů je méně omezené než působení adrenalinu a pokrývá mnoho oblastí mozku. Působení acetylcholinu na retikulární formaci je opačné než jeho periferní účinek na vnitřní orgány. Retikulární formace střední a prodloužené míchy excituje oxid uhličitý.
Hormony a mediátory působí na funkci mozkových hemisfér jak přímo, tak prostřednictvím retikulární formace. Retikulární formace mozkového kmene je tedy subkortikálním centrem autonomního nervového systému.
otázka 2.
- množství jaderných struktur, které zaujímají centrální místo v mozkovém kmeni. Morfologická stavba retikulární formace velmi připomíná síťovinu a právě na tuto vlastnost poprvé upozornil německý anatom Otto Deiters. Právě na tomto základě pojmenoval tuto strukturu (lat. reticulum – pletivo, formatio – vzdělání). Souhlasím! „Mesh education“ nezní tak cool 🙂
Vzhledem k tomu, že retikulární útvar prochází celým mozkovým kmenem, anatomicky by se dal rozdělit na úseky prodloužené míchy, pons varolii a středního mozku, ale protože jednotlivé části této struktury jsou pro ně obsazeny společným úkolem, považuje se za jediná struktura.
Pro pochopení uvedu srovnání ... Pokud jste někdy byli na koncertech nebo je alespoň sledovali v televizi, pravděpodobně jste si všimli obrovských panelů s hromadou tlačítek, páčkových přepínačů, přepínačů atd. Pomocí těchto panelů operátor upravuje kvalitu zvuku tlumením některých frekvencí a zvýšením jiných, stejně jako barvu, jas, kontrast atd. Takže retikulární formace dělá právě to. To znamená, že přijímá signály z absolutně všech sestupných a vzestupných cest, zpracovává je, vyvíjí nové koordinační signály a vydává je pro zamýšlený účel, čímž nám poskytuje normální vnímání.
Některé neurony R.t. vykazují aktivitu na pozadí, vybíjejí se 5-10krát za sekundu. Tato nervová centra ovlivňují mozkovou kůru a neustále v nás udržují vědomí. Se zničením těchto center u zvířat dochází.
Dovolte mi vysvětlit pro srovnání. Retikulární formace nás udržuje při vědomí stejným způsobem, jako oheň v ohni udržuje vodu v kotli vařící. Jakmile je oheň uhašen, vroucí voda, stejně jako vědomí, přestává být aktivní. V důsledku toho v R. f. jsou jedním z center spánku a bdění.
S mozkovou kůrou obecně jsou spojení zvláštní. Je jasné, že R.F. zodpovědný za a excitaci v mozkové kůře. Mozková kůra má zase také inhibiční a excitační účinky na retikulární formaci. Tyto dva systémy tvoří uzavřená neurální spojení, vzájemně se regulují a vyrovnávají své vlivy.
závěry
Funkce retikulární formace nebyly dostatečně prozkoumány kvůli vysokému stupni složitosti organizace této struktury, ale dostupná data jsou dostatečná pro následující závěry:
Ovlivňuje úroveň vědomí interakcí s mozkovou kůrou. Účastní se cyklu v dávání emocionální zbarvení smyslové signály, včetně bolesti, vedením aferentní informace do limbického systému. Vzájemnou koordinací aferentních a eferentních systémů se podílí na tvorbě vitálních reflexů. Podílí se také na autonomních funkcích těla a jako důležitá součást motorických center mozkového kmene.
Složitá struktura lidského mozku odhaluje tajemství našeho chování, vysvětluje zákonitosti duševní aktivita proud emocí a pocitů. Každá hemisféra mozku je zodpovědná za své specifické funkce a úkoly (např. je známo, že pravá je zodpovědná za logiku a levá za představivost a fantazii), ale existují i struktury, které zajišťují jednotnou a koordinovanou práci celého centrálního nervového systému. Jednou z těchto struktur je retikulární formace.
Obecná informace
Retikulární formace je úsek mozkového kmene, reprezentovaný rozsáhlou sítí nervových buněk a jader, které spojují různé části mozku. Na rozdíl od jiných struktur - například thalamus, hypothalamus, mozeček - které mají určitý celistvý tvar (jádra, žlázy), retikulární formace není reprezentována jediným morfologickým útvarem, ale je "sítí" (z latinského reticulum - síť) dendritů a axonů , které s různé míry hustoty pronikají mezi útvary a struktury mozku, vzájemně je spojují a zajišťují jejich společnou činnost.
Metaforicky řečeno: pokud je náš mozek prezentován ve formě určitého produktu, řekněme košile, pak retikulární útvar jsou nitě, kterými je košile šitá. Retikulární formace prostupuje strukturami prodloužené míchy, středního mozku a mostu, má přímé spojení s mozečkem, míchou, thalamem a nepřímé spojení s nadložními úseky: hypotalamem, zrakovými jádry a kůrou.
Jak to funguje
Retikulární útvar obsahuje velké množství neurony s rozvětvenými dendrity a dlouhými axony, díky nimž je možné přenášet nervové impulsy do různých částí mozku a míchy. V tomto případě lze rozlišit dvě největší skupiny neuronových shluků:
- Retikulo-tegmentální jádro, jehož neurony přijímají signály z nadložních částí GM (quadremium, thalamus) a přenášejí je dále do struktur mozečku, čímž regulují některé životně důležité motorické funkce: koordinaci pohledu, pohyby očí.
- Na regulaci dýchání a cévní inervace se podílí laterální jádro, jehož neurony vystupují ze struktur míšních a vestibulárních jader a poskytují GM kortexu informace o poloze těla v prostoru.
- Retikulární formace navíc zahrnuje neurony, které jsou důležité v práci center termoregulace, sytosti a hladu.

Hlavní funkce
Hlavním účelem retikulární formace je senzorická analýza četných signálů přicházejících z různých částí GM.
Díky úzkému spojení s míchou se také aktivně podílí na motorické regulaci, od polykacího reflexu až po složité motorické operace. Retikulární formace má navíc aktivační účinek na celý GM jako celek, podílí se na regulaci cyklů spánku a bdění.
V obecný pohled funkce retikulární formace jsou následující:
- Regulace kosterního svalstva (podílí se na řízení pohybů těla) a autonomních funkcí (dýchání, kýchání, krevní oběh atd.).
- Řízení procesů spánku a bdění (poskytnutím aktivačního a inhibičního účinku na mozkovou kůru).
- Aktivační funkce (projevuje se tím, že retikulární formace zajišťuje neustálou tonickou excitaci mozkové kůry, díky čemuž je možné udržet pozornost, vědomí a tok myšlenkových procesů.)
- Zpracování signálů přicházejících z vnějšího a vnitřního prostředí.
Charakteristické rysy práce retikulární formace jsou spojeny především s jejími určitými vlastnostmi:
Nervové buňky, které tvoří retikulární formaci, mají zvýšenou schopnost tonické excitace. To znamená, že většina neuronů je v neustálé excitaci a generuje nervové impulsy, které jsou přenášeny do překrývajících se částí GM. Tato tonická aktivita je způsobena řadou faktorů:
- Propustnost velkého množství signálů strukturami retikulární formace. Pro jednoduchou analogii si představte klavír nebo jiný strunný nástroj. Je jasné, že když se struny přímo dotkneme, začnou vibrovat a vydávat zvuk. Totéž se děje s nervovými buňkami, když k nim přicházejí signály z jiných neuronů. Představme si však dále, že se nedotýkáme přímo strun nástroje, ale řekněme vedle něj skáčeme a silně bušíme nohama o podlahu. Zvuk nástroje sice neuslyšíme, ale i tak bude probíhat sotva znatelné chvění strun. Totéž se děje s neurony retikulární formace. Protože jím neustále procházejí některé signály (aferentní i eferentní) z různých struktur centrálního nervového systému, vytváří to neustálou tonickou excitaci neuronů retikulární formace, protože je v epicentru konstantního nervu. - výměna impulsů.
- Zvýšená citlivost neuronů na Chemikálie(hormony, drogy, psychotropní látky). Ráno vypitý šálek kávy „zapíná“ přesně struktury retikulární formace a díky dlouhodobému uchování vzruchu v jejích neuronech nás udržuje v aktivním stavu.
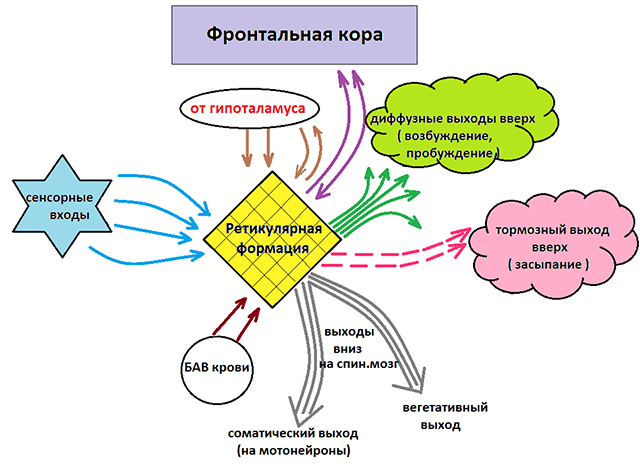
Sestupný a vzestupný vliv Ruské federace
Jak již bylo uvedeno, retikulární formace má excitační a inhibiční účinek na různé části GM. V tomto případě lze rozlišit dvě oddělení, která se specializují na přenos vzruchu do určitých mozkových struktur.
Sestupné oddělení: zastoupeno vegetativními a motorickými centry a působí směrem dolů na míchu. Odpovídající nervové akumulace regulují činnost respiračních, vazomotorických, slinných center a také center odpovědných za budování jednoduchých a složitých motorických reakcí. To ukazuje na rozhodující roli centrálního nervového systému v regulaci i elementárních nepodmíněných reflexů. Stimulace sestupného úseku vede k inhibici páteřních center a vyvolává stav hlubokého spánku v přirozeném prostředí (spánek „bez zadních nohou“). Stejný efekt lze vyvolat i uměle, například uvedením člověka do stavu transu nebo anestezie.
Ascendentní oddělení: představováno nervovými vlákny spojujícími struktury retikulární formace s nadlehlými odděleními: thalamus, hypotalamus, mozeček a kůra. Vzestupný vliv působí stimulačně na korové struktury a zajišťuje aktivní stav vědomí. Vzestupný vliv neustává ani když spíme. Kdyby se náš mozek dokázal úplně „vypnout“, každé probuzení by se podobalo narození: kdo jsem? Kde jsem? Jak jsem se sem dostal? Díky práci retikulárních struktur si však zachováváme možnost vždy se vrátit do výchozího stavu vědomí, ve kterém jsme byli před okamžikem spánku. Navíc během nočního klidu máme stále schopnost reagovat na určité životně důležité podněty, tzn. zpravidla nespíme v „mrtvém spánku“ a můžeme se probudit, pokud se dítě pohybuje a pláče poblíž, něco hlasitě spadne atd.
Projev poškození konstrukce
Retikulární formace hraje významnou roli v integrační aktivitě celého mozku. Vzhledem k funkci vedoucího vodiče všech typů nervových vzruchů do všech částí centrálního nervového systému je retikulární formace v neustálém provozu. Nadměrné duševní a emoční přetížení je škodlivé pro mozek obecně a pro retikulární formaci zvláště. Naštěstí včasné přijetí sedativa může (vzhledem ke zvýšené náchylnosti neuronů k chemické expozici) rychle napravit situaci a normalizovat stav.
Jsou však možné i méně příznivé výsledky. Poškození je možné v důsledku traumatického poranění mozku, onkologických onemocnění mozku, infekčních lézí.
Hlavním projevem potíží je ztráta vědomí.
Porušení vzestupných spojení se projevuje ve stavu apatie, slabosti, zvýšené ospalosti, motorické disinhibice, poruch nočního spánku. Neobvyklé nejsou ani doprovodné vegetativní poruchy.
Retikulární formace (latinsky rete - síť) je soubor buněk, buněčných shluků a nervových vláken umístěných v celém mozkovém kmeni (medulla oblongata, most, střední mozek a diencephalon) a v centrálních částech míchy. Retikulární formace přijímá informace ze všech smyslových orgánů, vnitřních i jiných orgánů, vyhodnocuje je, filtruje a přenáší do limbického systému a kůry velký mozek. Reguluje úroveň dráždivosti a tonusu různých částí centrálního nervového systému včetně mozkové kůry, hraje důležitou roli ve vědomí, myšlení, paměti, vnímání, emocích, spánku, bdění, autonomních funkcích, cílených pohybech a také v mechanismech tvorby integrálních reakcí těla. Retikulární formace primárně funguje jako filtr, který umožňuje důležitým senzorickým signálům pro tělo aktivovat mozkovou kůru, ale nepropouští navyklé nebo opakující se signály.
Retikulární formace je důležitým bodem na cestě vzestupného nespecifického somatosenzorického systému. Somatoviscerální aferenty jdou jako součást spinoretikulárního traktu (anterolaterální funiculus), případně také jako součást propriospinálních (polysynaptických) drah a odpovídajících drah z jádra spinálního trigeminálního traktu. Do retikulární formace přicházejí i cesty ze všech ostatních aferentních hlavových nervů, tzn. téměř ze všech smyslů. Další aferentace pochází z mnoha dalších částí mozku – z motorických oblastí kůry a senzorických oblastí kůry, z thalamu a hypotalamu. Existuje také mnoho eferentních spojení - sestupných do mícha a vzestupně přes nespecifická thalamická jádra do mozkové kůry, hypotalamu a limbického systému. Většina neuronů tvoří synapse se dvěma nebo třemi aferenty různého původu, taková polysenzorická konvergence je typická pro neurony retikulární formace. Jejich dalšími vlastnostmi jsou velká receptivní pole povrchu těla, často oboustranná, dlouhá latentní doba odezvy na periferní stimulaci (díky multisynaptickému vedení), špatná reprodukovatelnost reakce (stochastické kolísání počtu akčních potenciálů při opakované stimulaci). Všechny tyto vlastnosti jsou opačné než vlastnosti lemniskálních neuronů ve specifických jádrech somatosenzorického systému (obr. 9-7 a obr. 5-13).
Funkce retikulární formace nebyly plně prozkoumány. Předpokládá se, že je zapojena do následujících procesů:
1. v regulaci úrovně vědomí ovlivňováním aktivity kortikálních neuronů, např. účastí v cyklu spánek/bdění,
2. při poskytování afektivně-emocionálního zabarvení smyslových podnětů, včetně signálů bolesti jdoucích podél anterolaterálního funiculu, vedením aferentní informace do limbického systému,
3. v autonomních regulačních funkcích, včetně mnoha životně důležitých reflexů (oběhové reflexy a respirační reflexy, reflexní akty polykání, kašlání, kýchání), při kterých musí být vzájemně koordinovány různé aferentní a eferentní systémy,
4. v účelných pohybech jako důležitá součást motorických center mozkového kmene.

Otázka 48. Srovnávací charakteristiky kabelového a salatorního buzení
Nervová tkáň má takovou fyziologickou vlastnost, jako je vodivost, tedy schopnost vést vzruch podél průběhu nervového vlákna ve formě akčního potenciálu. Existují dva typy vedení vzruchu v závislosti na struktuře nervového vlákna. Existují dva typy nervových vláken: dužinatá (myelinizovaná) a amyelinizovaná (nemyelinizovaná). U nemasitých nervových vláken je pozorováno kontinuální šíření vzruchu, které je založeno na lokálních nebo kruhových proudech. Jak již bylo zmíněno dříve, excitovaný elektronegativní úsek nervového vlákna se stává dráždivým pro nejbližší neexcitovaný elektropozitivní úsek, který je excitován (excitovaný úsek je jakoby vybíjen směrem k neexcitovanému, což má za následek vznik lokálního nebo kruhového proudy).
Myelin, pokrývající nervové vlákno, se nachází v segmentech, tedy nespojitě. Myelin je dobrý izolant a kdyby pokryl nervové vlákno souvislou vrstvou, tak by se vzruch nerozšířil. Myelinová pochva je tvořena buňkami neurilemy nebo Schwannovými buňkami. Plazmatická membrána jedné Schwannovy buňky spirálovitě obaluje několika vrstvami část axonu o délce několika setin mikronu. Mezi úseky axonu pokrytými myelinovou pochvou zůstávají nemyelinizované zóny. Tyto zóny se nazývají záchyty Ranviera.
Ve vláknech pokrytých myelinovou pochvou (pulpová vlákna) se vzruch šíří křečovitě (saltatoricky), tedy podél Ranvierových záchytů. Jak ukázal japonský fyziolog Tasaki, vytváří se tak jakýsi systém spolehlivosti pro šíření vzruchu (potenciální rozdíl mezi excitovanými a nebuzenými úseky vlákna stačí na 5-6 zachycení Ranviera). V případě, že dojde vlivem velkého elektrického pole k poškození na malém úseku vlákna, nedochází k narušení šíření akčního potenciálu. Jak je známo, počáteční část axonu v nervové buňce není pokryta myelinovou pochvou. Právě v tomto úseku vlákna se tvoří akční potenciál. Existuje potenciální rozdíl mezi vybuzeným a nevybuzeným úsekem prvního zachycení Ranviera, který je pod vlivem tohoto elektrického pole vybuzen. Poté se vytvoří potenciální rozdíl mezi excitovaným prvním Ranvierovým uzlem a následujícím, který se dobíjí a excitace získává propagační charakter. Základem pro šíření vzruchu po dužinatém vláknu, ale i nemasitém vláknu, jsou tedy také lokální (kruhové, vířivé) proudy. V uzlech Ranvier, umístěných ve vzdálenosti 2 mm od sebe, byla nalezena vysoká hustota sodíkových kanálů - až 1200 na 1 μm2, což značně usnadňuje vedení vzruchu podél nervového vlákna. Diskontinuální šíření buzení má některé výhody oproti kontinuálnímu šíření. Za prvé, rychlost šíření vzruchu u myelinem potažených vláken je 8-10krát rychlejší než u amyelinizovaných. Za druhé, na šíření excitace nespojitého typu je vynaloženo méně energie, je to ekonomičtější, což se vší pravděpodobností souvisí s vysokou hustotou sodíkových kanálů v Ranvierových uzlech.
Když se vzruch šíří podél nervového vlákna, je třeba vzít v úvahu čistě fyzikální nebo kabelové vlastnosti vodiče (nerv si lze představit jako kabel umístěný v mořské vodě). Mezi vlastnosti kabelu patří zejména průměr (průřez) vodiče - čím silnější je nervové vlákno (nebo čím větší průřez), tím nižší je odpor. Tím rychlejší bude tedy šíření vzruchu ve formě impulsu. Velký význam při buzení má také kapacita a odpor membrány. Pokud je tedy vstupní odpor membrány větší, pak se vzrušivost v tomto místě snižuje. Mezi vlastnosti kabelu patří také elektrický tón, který má velký vliv na vodivost: čím výraznější katelectroton, tím rychleji je veden akční potenciál. Anelektrotonické změny naopak zhoršují vedení vzruchu nervovou tkání.
Podle rychlosti vedení vzruchu se všechna nervová vlákna dělí do tří skupin: A, B a C. Nervová vlákna skupiny A jsou vysokorychlostní vlákna výhradně dužnatého typu. V závislosti na průřezu nervového vlákna se rychlost jejich excitace pohybuje mezi 20-120 m/s. Existují A-vlákna - nejrychlejší - 70-120 m/s (průměr vlákna 12-20 mikronů - a-vlákna, jejich průměrná rychlost buzení je 70-120 m/s; průměr 8-12 mikronů - b-vlákna, vodivá buzení rychlostí 40-70 m/s, průměr vlákna 4-8 mikronů - g-vlákna provádějící buzení rychlostí 20-40 m/s). Čím silnější je vodič, tím větší je rychlost buzení. Nervová vlákna skupiny B jsou převážně nemasitá vlákna, rychlost šíření vzruchu je 6-20 m/s. Nervová vlákna skupiny C jsou zastoupena výhradně nemasitými vlákny vegetativní povahy, rychlost jejich excitace je 0,5-6 m/s.
Ve fyziologii existují tři zákony šíření vzruchu.
Zákon celistvosti nervu (zákon kontinuity). Nerv vede excitaci pouze tehdy, pokud si zachová svou histologickou a funkční integritu. Jakékoli odchylky těchto indikátorů vedou k porušení jeho vodivosti. Působení lokálních anestetik (novokain) je založeno na skutečnosti, že molekuly novokainu blokují sodíkové kanály, v důsledku čehož se sodíkový proud zastaví a tkáň ztrácí schopnost excitace. Vzrušení při dráždění receptorů bolesti dosáhne místa, kde působí novokain, a je zablokováno, v důsledku čehož se impulsy bolesti nedostanou do centra bolesti.
Zákon oboustranného buzení. Nervové vlákno je schopno vést vzruch z receptorů do center a naopak, z center do periferních útvarů. Takový vzor byl prokázán v klasických studiích Kuehne a Babukhin. Kuehneova zkušenost tedy byla následující: pokud je narušena integrita svalu mezi jeho dvěma sekcemi, které jsou inervovány dvěma větvemi jednoho axonu, pak elektrická stimulace kterékoli z větví axonu vede ke kontrakci obou částí. svalu.
Zákon izolovaného šíření vzruchu. Je známo, že akční potenciál ve vláknech pokrytých myelinem se nepřenáší z jednoho nervového vlákna na druhé díky dobrým izolačním vlastnostem myelinu. Takové izolované vedení buzení zajišťuje malé a přesné profesionální svalové kontrakce (hra na klavír, práce hodináře atd.). Bezprostředně po narození nedochází k dostatečné myelinizaci nervových vláken a novorozenci na jakékoli podráždění ve většině případů reagují nikoli lokálními, ale difúzními kontrakcemi velké svalové skupiny. Podobná reakce je pozorována u všech hladkých svalů, které jsou inervovány nemasitými nervovými vlákny, která nemají izolační vlastnosti.